
From within-host interactions to epidemiological competition: a general model for multiple infections
While much of our epidemiological theory has been developed for single pathogen – host systems, a growing body of research is finding that multiple parasite strains or species often infect the same host in nature. The interactions between different strains of pathogens are not trivial and can vary from symbiotic to antagonistic. The unpredictable outcome of the within-host interactions between parasites and the possible combinations of co-transmission are two reasons the authors suggest that the majority of epidemiological theory and models are lacking.
To address this gap in research, the authors developed a model for multiple infections which incorporates differential outcomes of interactions between parasites, a variable number of parasite genotypes, and the possibility for any combination of co-transmissions between each available parasite genotype within the host.
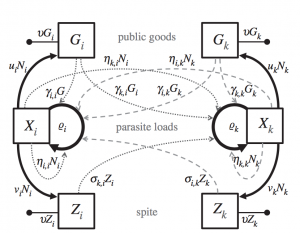
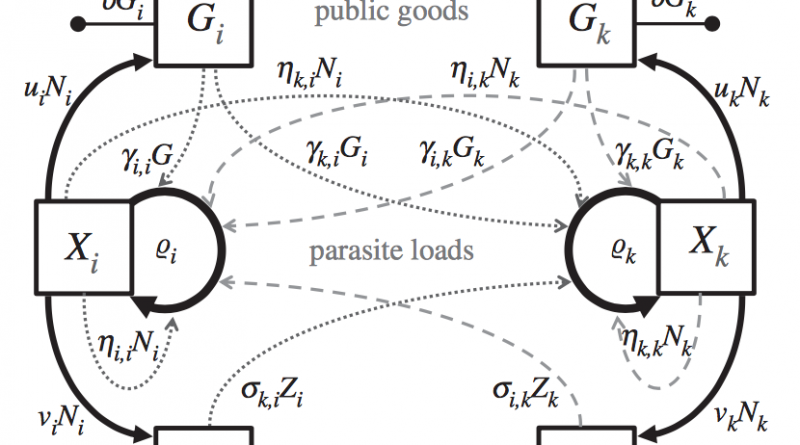
They modeled outcomes of interactions between parasites by modeling public production dynamics as either “public goods” or “spite”. Public goods are resources that increase receiver growth rate (e.g., siderophores). Spite are public productions decrease receiver growth rate (e.g., bacteriocins). Their model assumed each genotype can produce either goods or spite but the particular good can have different effects on the receiver depending on their genotype. To incorporate this they defined a matrix of goods and a matrix spites which determine the effect of the production on each genotype. In addition to goods and spite, they also incorporated a density-dependence matrix which determines the effect of growth rates of each genotype on the growth rate of other genotypes. The density-dependent matrix could represent effects such as competition or indirect interaction through elicitation of the immune response. Finally, they made a vector of parasite growth rates for each genotype.
They analytically calculate the dynamics of this model by assuming that dynamics of parasite load are at steady state. They imposed time-scale separation of the within and between host dynamics to make inferences at the between host level. The between host system is an extended SIS model with sequential recovery in a host population of constant size. Susceptible individuals can immediately become co-infected by a co-infected individual. Recovery in their model only happens for one genotype at a time. They calculated R0 using the infected steady state density of hosts and found that R0 can show complex behaviors when epidemiological parameters vary (i.e., R0 was a nonlinear function of β). They suggested that this could lead to public health interventions that aim to decrease β but only end up increasing R0.
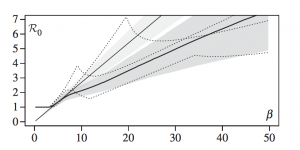
They suggested that their model could be used for parameter inference for diseases with detailed epidemiological data available (e.g., HPV). Overall, this paper was interesting for this class because it was one of the first to tackle several within-host interactions (e.g., spite, goods, and density-dependence) for more than two parasite genotypes. They showed how R0 was a non-linear function of transmission such that decreasing transmission (β) could increase spread within a population.